
Proprioception: You’re Looking at the Wrong End of the Foot
Authored by Sam Fowler DipWCF & The Hoofflix Team
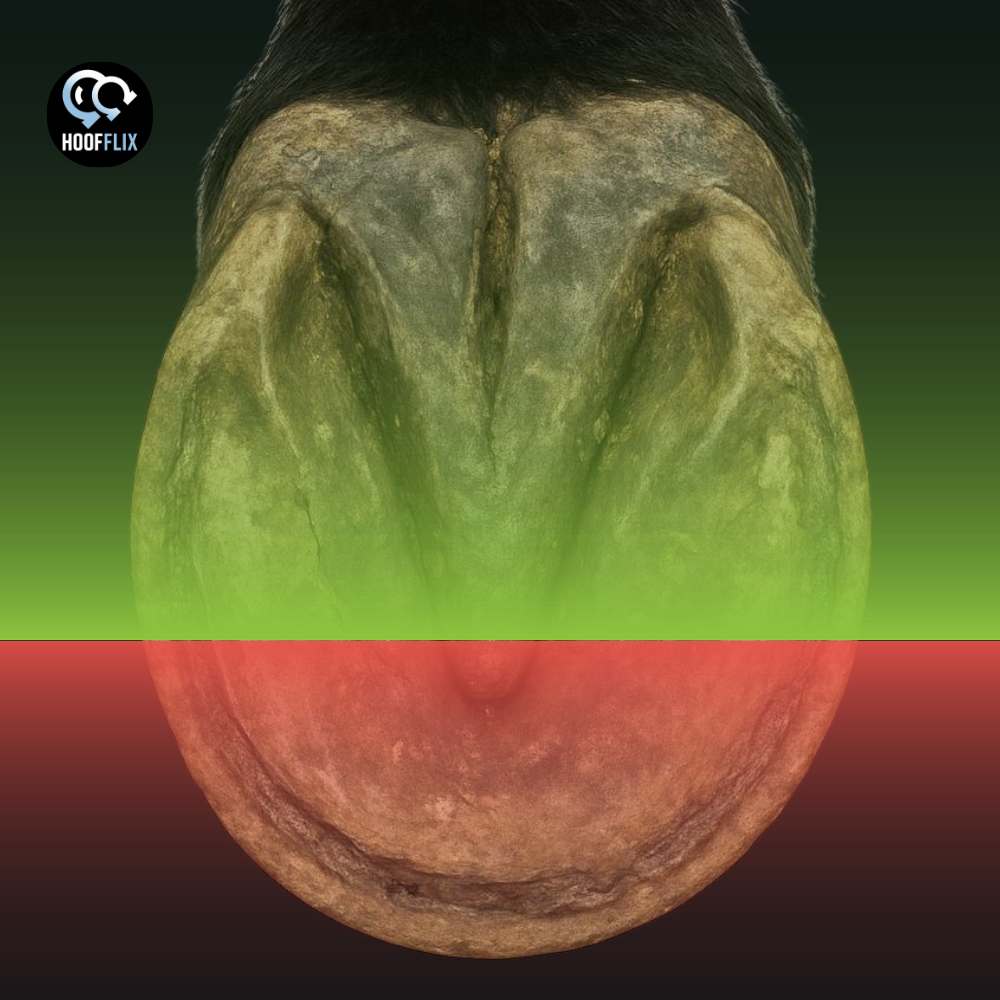
The proprioceptive receptors in the equine foot are concentrated in the frog and palmar heel. The toe does not contain them. The online conversation has been arguing about the wrong structure, and it is not the first time.
Before we start: what proprioception actually is
The word “proprioception” gets used loosely online, often as a synonym for “feeling” or “sensation.” It is not. The foot has several distinct sensory systems, and they do different jobs.
Proprioception is the sense of where your body is in space and how it is moving. Close your eyes and touch your nose: that is proprioception. It tells the nervous system the position and movement of limbs, joints, and load-bearing structures without needing to look at them. In the horse’s foot, proprioceptors detect ground impact, changes in loading, and shifts in limb position, feeding that information into reflexes that adjust stance and gait in real time. It is an unconscious system. The horse does not “feel” it the way it feels pain; the nervous system uses it automatically.
Nociception is pain sensing. A different set of nerve fibres, using different receptor types, carrying signals at different speeds. When a horse steps on a sharp stone, that is nociception. It is a warning system: something is wrong, move away from it. Nociceptive fibres are predominantly unmyelinated and slow. Proprioceptive fibres are myelinated and fast. They are not the same network.
Vascular innervation is a third system: nerves that regulate blood flow through the foot’s microvasculature. These use chemical messengers (substance P, neurokinin A) to dilate or constrict blood vessels. They keep the foot perfused. They do not tell the horse where its foot is.
These three systems coexist in the foot, but they are anatomically distinct. Different receptor types, different fibre types, different locations. The dorsal wall and toe are not “nerveless”: they have nociceptive fibres and vascular innervation. But none of that nerve supply can sense the length of the toe, detect the mechanical consequences of a long lever arm, or tell the horse where breakover is happening. The toe’s innervation can detect tissue damage and regulate blood flow. It cannot detect position, pressure, vibration, or ground texture. The critical question for the proprioception argument is not “does the toe have nerves?” (it does) but “does the toe have proprioceptive nerves?” That distinction is what the rest of this article is about.
You will have heard the argument: the horse’s foot is a sensory organ; shortening the toe or applying a shoe destroys proprioception; the foot needs to feel the ground. Parts of that are true. The foot is a sensory organ. Proprioception matters. But the argument has a fatal anatomical problem, and it needs to be stated plainly before anything else.
Here is the crux. The proprioceptive receptors in the equine foot are not distributed evenly across the sole. They are overwhelmingly concentrated in the palmar (caudal) foot: the frog, the heels, the digital cushion, and the tissues surrounding the navicular bone. The dorsal hoof wall and toe are innervated, but their innervation is nociceptive (pain-sensing) and vascular, not proprioceptive. The claim that shortening the toe “destroys proprioception” requires the toe to contain proprioceptive apparatus. It does not. That is not an opinion. It is the consistent finding of every published receptor-distribution study conducted on the equine foot.
In a laminitic horse, the argument becomes even weaker because the wall at the toe is connected to the coffin bone by a laminar wedge of scar tissue that cannot transmit any sensory signal at all. The structure being “protected” has no sensory function and is connected to nothing functional. Meanwhile, the lever arm it creates is overloading the deep flexor tendon at every stride.
This article lays out the evidence, connects it to the arguments in the earlier HoofFlix articles Two Levers, One Hoof and What “Just Drop The Heels” Isn’t Telling You About Your Laminitic Horse, and explains what genuinely proprioceptive management of the foot actually looks like.
The evidence: where the receptors sit
Bowker and colleagues published the foundational receptor-distribution study in 1993.1 Lamellated corpuscles with the morphological characteristics of Pacinian corpuscles were identified in the hoof dermis, but they were restricted to the palmar (caudal) aspects of the solar dermis of the heel and not scattered through the foot and concentrated in the back half, beneath and around the frog and heels.
Pacinian corpuscles are rapidly adapting receptors. They fire in response to abrupt pressure changes and vibration: exactly the stimulus generated when a heel-first landing strikes the ground. They give the nervous system a positional update at each stride, allowing the horse to adjust reflexes and muscle activity for the next step.1,2
The study also identified naked nerve endings containing calcitonin gene-related peptide (CGRP) in the skin, solar dermal tubules, and digital cushion.1 These serve nociceptive and vasomotor functions: pain and blood-flow regulation, not proprioception.Bowker’s 1995 follow-up examined sensory nerve fibres across the equine distal forelimb.2
Sensory peptides innervated much of the foot, including the dermis of the dorsal wall. But the nerve fibres supplying the foot were predominantly unmyelinated over myelinated (approximately 3.7 to 1).2 Unmyelinated fibres carry slow nociceptive signals and autonomic vascular control. Myelinated fibres carry fast proprioceptive information. The dorsal wall has nerve supply. It is not proprioceptive nerve supply.
Van Wulfen and Bowker confirmed this in 2002 by specifically examining the dorsal hoof wall for tachykinins (substance P and neurokinin A).3 They found tachykinin receptors on the microvasculature: a vasodilatory mechanism for blood-flow control. No proprioceptive receptor structures were identified in the dorsal wall.
Castelijns in 2017 looked for Merkel cells and small lamellated Pacinian-like corpuscles (SLPCs) in both the dorsal wall and the frog.4 Merkel cells (slowly adapting touch receptors) were at the dermal-epidermal junction of the frog. SLPCs were in the frog dermis, associated with the central ridge. The proprioceptive and touch-receptor apparatus was, once again, in the frog.
The pattern across three decades of published work is consistent. The proprioceptive system lives in the caudal foot. The dorsal wall and toe have vascular and pain innervation. These are different neurological systems doing different jobs. A note on the research itself. Bowker’s work is frequently cited within the barefoot and alternative hoof care communities as evidence that the foot is a sensory organ that must not be interfered with. His work does show the foot is a sensory organ. But his own findings place the proprioceptive apparatus in the frog and palmar heel, not in the dorsal wall or toe. The barefoot community has been citing his research correctly (“the foot is a sensory organ”) and then misapplying it to the wrong part of the foot (“therefore do not touch the toe”). Bowker’s work on conformable surfaces, substance P release, and frog engagement is about the caudal foot doing its job. It has never been about the toe.
It is also worth addressing the claim that the dorsal wall might contain proprioceptive receptors that have simply not been found yet. The absence-of-evidence argument does not hold here.
Three decades of published research, using histology, immunocytochemistry, electron microscopy, and immunohistochemistry, have specifically examined the dorsal hoof wall.1,2,3,4
Multiple research groups, multiple methodologies, multiple sample sites. Every study that looked for mechanoreceptors in the dorsal wall came back with the same finding: vascular and nociceptive innervation, no proprioceptive structures. That is not a gap in the research. It is a consistent result.
The lamellae and the laminar wedge
The online proprioception argument often implies a sensory chain: ground contact at the toe transmits a signal through the wall, through the lamellae, to P3, and the horse “feels” the ground through this pathway. This treats the lamellae as a signal conduit. They are not. They are a suspension systems.
The lamellae transmit load from the skeleton through P3 to the hoof wall. Their nerve supply is vascular and nociceptive.2,3 They regulate their own blood supply and generate pain signals when damaged. They do not contain the mechanoreceptor apparatus (Pacinian corpuscles,
Merkel cells, Ruffini endings) required for proprioception.
The rivets holding a car door to its hinges transmit the weight of the door. They do not transmit the vibration of the road to the driver’s hands. The lamellae are structural. They bear load, not sensation.
In a laminitic foot, this becomes even more decisive. When the lamellar bond fails, the space between P3 and the dorsal wall fills with pathological tissue: the laminar wedge. This is keratinised scar tissue, a wound-healing response producing ectopic white line composed of hyperplastic and hyperkeratotic epidermal tissue.6 It is structurally inferior to the original lamellae, mechanically unreliable, and incapable of transmitting the rapid, precise mechanical stimulus that proprioceptors require.
The fingernail analogy from Two Levers, One Hoof extends naturally here. A fingernail partly lifted from the finger, with scar tissue filling the gap, does not “feel” the table surface when you press it down. Press on the nail and you feel pressure at the nail bed where it is still attached; you feel nothing useful through the scar tissue. The interface is dead. The same applies to the laminar wedge. The wall at the toe looks like hoof wall, but it is anchored to scar tissue, not to a functional lamellar bond, and even that functional bond was never part of the proprioceptive apparatus.
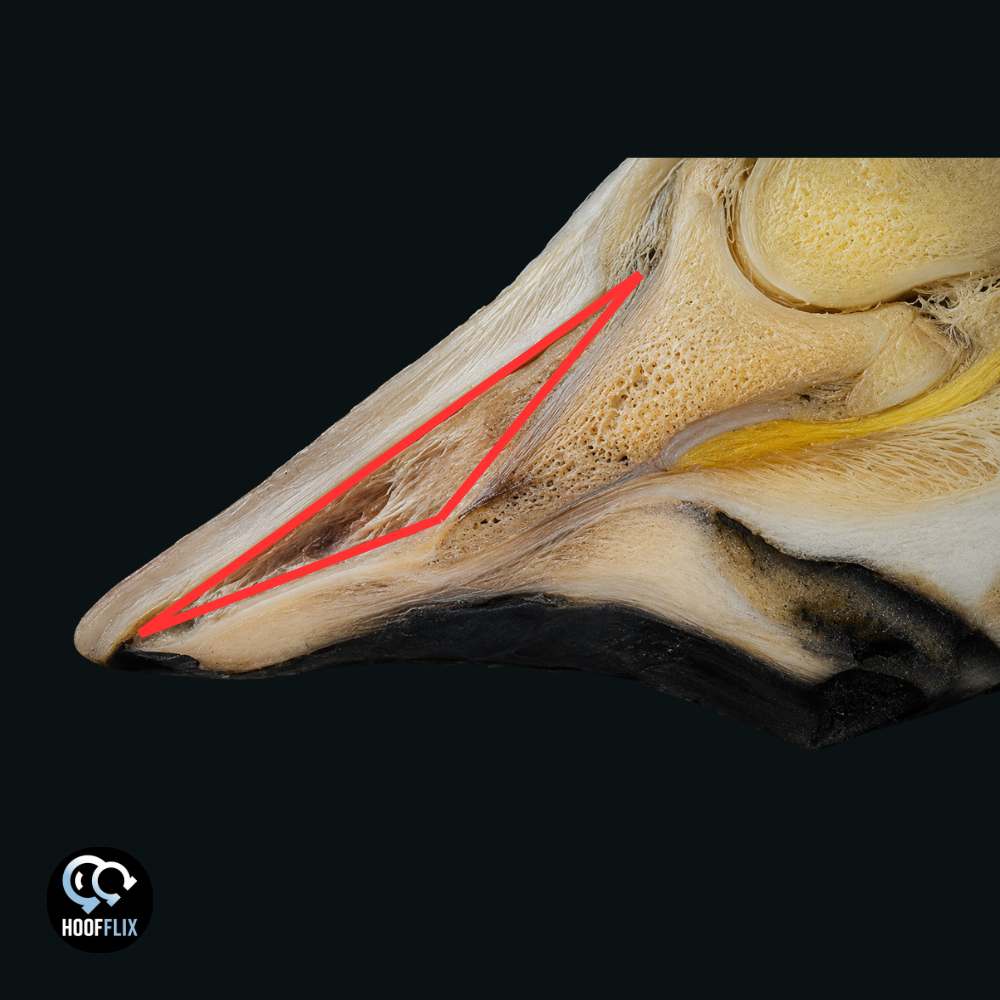
A related argument circulates under different terminology: the toe wall is a “balance point” or “pillar,” and shortening it causes the capsule to rotate, P3 to shift, and the horse to be crippled. The claim runs cause and effect backwards. In a laminitic foot, P3 has already rotated because the lamellar bond failed and the deep flexor tendon pulled the bone. The toe wall is not holding P3 in place; the laminar wedge has already disconnected it. A pillar requires a structural connection to the thing it supports, and the laminar wedge provides none. Shortening that disconnected wall does not cause rotation. Rotation already happened. What shortening does is reduce the lever arm that is driving further mechanical damage at every stride. The claim that sole callousing after toe shortening is a “trauma response” mistakes normal weight-bearing adaptation for pathology; the sole is doing the job it was designed to do, which is to bear load, on a foot whose wall was never bearing it properly because the lamellar bond was already gone.
The argument that shortening the toe “destroys proprioception” requires two things to be true that are both false: that the dorsal wall contains proprioceptive receptors, and that the laminar wedge can transmit proprioceptive signals.
A common counterargument runs as follows: even if the toe itself has no proprioceptors, when the toe contacts the ground the whole hoof capsule deforms, and the caudal receptors sense that deformation. This is partially true: the capsule does deform under load, and the caudal receptors do detect deformation. But a long toe does not improve that signal. It distorts it. The deformation pattern generated by a long lever arm is not the same as the deformation pattern of a balanced foot landing heel-first on a conformable surface. A long toe generates asymmetric, excessive strain concentrated through the dorsal wall and the DDFT; the caudal receptors are not receiving a richer proprioceptive input; they are receiving a louder mechanical overload signal. More deformation is not more proprioception. It is more strain. There is a further problem. The proprioceptive receptors in the caudal foot can detect the mechanical consequences of a long toe, but what they detect is not something worth preserving. The Pacinian corpuscles clustered around the DDFT and navicular ligaments, and the Ruffini endings in the neurovascular bundles and sole, sit directly in the path of the forces a long toe generates. A long lever arm means increased DDFT tension, increased navicular compression, increased strain through the very structures these receptors are embedded in.
The proprioceptive system is not telling the horse “my toe is the right length.” It is telling the horse “there is too much load coming through here.” Shortening the toe reduces that load. The caudal proprioceptors register a 10 mm breakover shift as a 21 per cent reduction in peak DDFT tension.7,8 That is the proprioceptive system working better, not being destroyed.
Preserving a long toe in the name of proprioception is preserving the stimulus that the proprioceptive system is warning the horse about.
What this means for farriery
Two Levers, One Hoof established that the toe functions as a lever arm. A 10 mm shift in breakover reduces peak deep digital flexor tendon tension by approximately 21 per cent.7,8 The farriery decision to shorten the toe is about reducing a mechanical lever that overloads the deep flexor system, not about removing sensation.
The heel-height article made the complementary point: the toe and heel are coupled through the bone. If the proprioceptive apparatus sits in the caudal foot, then heel height, frog engagement, and palmar angle are the variables that affect proprioceptive input. Toe length is not a proprioceptive variable. It is a mechanical one. Confusing the two leads to clinical decisions that protect the wrong structure for the wrong reason.
Three scenarios where the proprioception argument does harm
Scenario one: the laminitic foot with a distorted toe.
The owner is told not to let the farrier shorten the toe because it will “remove proprioception.” The lever arm stays long. The DDFT load stays elevated. The mechanical damage continues because the owner has been told the toe is a sensory organ rather than a lever.
Scenario two: the chronic foot with a laminar wedge.
The toe wall is separated from P3 by scar tissue. No functional lamellar bond exists at the toe. The material is contributing nothing structural and nothing sensory, but it is extending the lever arm and resisting breakover. The owner has been told to preserve it because “the foot needs to feel.” The material cannot feel anything.
Scenario three: the horse was kept from appropriate shoeing.
The owner is told shoes “destroy proprioception.” The horse is sore, landing toe-first, and developing palmar foot pain. The frog,where the proprioceptive receptors sit, is not being loaded because the horse is avoiding heel contact. The proprioception argument has produced the exact outcome it claims to prevent. It is worth being precise here: the claim that shoes impair proprioception is sometimes true, but not for the reason given. A shoe that lifts the frog entirely off the ground does reduce frog engagement, which does reduce proprioceptive input. But that is a shoe-fitting problem, not a fundamental property of shoeing. A well-fitted shoe with appropriate frog support, or a shoe with a pad, allows frog contact. The question is whether the frog is engaging the ground, not whether the horse is shod. And the question has nothing to do with the toe.
What good proprioceptive management looks like
Frog engagement is the priority, not toe preservation. The frog is where Pacinian corpuscles, Merkel cells, and substance P-releasing touch receptors are concentrated.1,4 Frog contact with the ground, whether barefoot or shod, is what activates the proprioceptive apparatus.
Heel-first landing is a proprioceptive intervention. The Pacinian corpuscles respond to rapid impact stimuli.1 Toe-first landing bypasses the proprioceptive apparatus entirely. Correcting the hoof balance to promote heel-first landing supports proprioception; preserving a long toe undermines it.
Conformable surfaces support the system. Bowker’s observations confirm that horses preferentially load the palmar foot on conformable surfaces, engage the frog, and exhibit reduced stress.2
Shortening the toe does not remove proprioceptive receptors, because there are none in the dorsal wall to remove. It reduces the lever arm, promotes heel-first landing, and improves the mechanical environment that allows the caudal foot to do its job. Toe management is a proprioceptive intervention, not an anti-proprioceptive one.
Questions worth asking
If “proprioception” is being used as a reason to preserve toe length or avoid farriery:
Where in the foot are the proprioceptive receptors?
A good answer names the frog and the palmar heel. An answer naming the whole foot or the wall is not consistent with the published studies.
Is the horse landing heel-first or toe-first?
Toe-first landing bypasses the proprioceptive apparatus. A management approach claiming to protect proprioception while permitting toe-first landing is contradicting itself.
Has the horse had radiographs showing the internal toe position?
In a laminitic horse, the external wall may be separated from P3 by scar tissue. You cannot see this from the outside.
Is the frog making contact with the ground?
Frog engagement is the primary activator of the proprioceptive system. If the frog is not touching the ground, proprioception is not being stimulated, regardless of toe length.
If the answers are “I don’t know,” whatever is being done to the feet is being done without the information needed to make it safely. That is not an attack. It is an invitation to ask for more, on behalf of a horse that cannot ask on their own.
The horses we worry about
A pony with chronic laminitis whose owner has been told not to let the farrier touch the toe because “the foot needs to feel.” The lever arm stays long. The DDFT stays overloaded. The laminar wedge, which is scar tissue connected to nothing functional, extends further forward while the owner believes it is a sensory structure worth protecting.
An owner who has been told shoes will “destroy proprioception” and is keeping a thin-soled horse barefoot on hard ground. The horse is landing toe-first to avoid heel pain. The frog is not being loaded. The proprioceptive system the owner is trying to protect is being bypassed entirely because the horse cannot use its heels.
A farrier who knows the toe needs to come back but faces an owner armed with social media posts about proprioception and unwilling to let the work be done. The farrier cannot explain in the time available why the argument is anatomically wrong. The toe stays long. The horse stays compromised. This article exists so that the conversation can be shorter.
A final word
The proprioceptive system in the equine foot is real. It just does not live where the online conversation says it does. The receptors are in the frog and palmar heel. The dorsal wall has pain and vascular innervation. The lamellae are a suspension system. The laminar wedge is scar tissue.
Share this with anyone whose horse is being managed under a protocol that uses “proprioception” as a reason to preserve toe length or avoid farriery intervention. The horses whose toes are being preserved for a sensory function that does not exist are the ones paying for the misunderstanding.
The receptors are in the frog. They always were. I just showed you where.
Where the numbers in this article come from
For owners who want to read further, or vets and farriers who want to check the references:
1. Bowker RM, Brewer AM, Vex KB, Guida LA, Linder KE, Sonea IM, Stinson AW. Sensory receptors in the equine foot. Am J Vet Res. 1993;54(11):1840-1844.
2. Bowker RM, Rockershouser SJ, Linder K, Vex KB, Sonea IM, Caron JP. Sensory nerve fibres and receptors in equine distal forelimbs and their potential roles in locomotion. Equine Vet J. 1995;27(Suppl 18):141-146.
3. Van Wulfen KK, Bowker RM. Evaluation of tachykinins and their receptors to determine sensory innervation in the dorsal hoof wall and insertion of the distal sesamoidean impar ligament and deep digital flexor tendon on the distal phalanx in healthy feet of horses. Am J Vet Res. 2002;63(2):222-228.
4. Castelijns H. Morphological evaluation of Merkel cells and small lamellated sensory receptors in the equine foot. Am J Vet Res. 2017;78(6):659-667.
5. Lancaster LS, Bowker RM, Mauer WA. Morphological and Biomechanical Properties of Equine Laminar Junction. J Equine Vet Sci. 2013;33(12):1002-1009.
6. Collins SN, van Eps AW, Pollitt CC, Kuwano A. The Lamellar Wedge. Vet Clin North Am Equine Pract. 2010;26(1):179-195.
7. van Heel MCV, Barneveld A, van Weeren PR, Back W. Dynamic pressure measurements for the detailed study of hoof balance: the effect of trimming. Equine Vet J. 2004;36(8):778-782.
8. Eliashar E, McGuigan MP, Wilson AM. Relationship of foot conformation and force applied to the navicular bone of sound horses at the trot. Equine Vet J. 2004;36(5):431-435.
9. Sharp Y. The Hoof-Horse Connection. The Equine Documentalist. 2021. Available at: https://www.theequinedocumentalist.com/the-hoof-horse-connection/